Selección genética en el galgo de campo

Introducción
En la medida que un carácter de interés, ya sea éste un carácter de comportamiento o una medida o carácter morfológico, está parcialmente condicionado por la acción de los genes es posible lograr una mejora utilizando una de las dos herramientas que tiene a su disposición un genetista: 1) la elección de los mejores reproductores dentro de la población, en este caso dentro de los galgos incluidos dentro del Libro Genealógico de la FEG, y; 2) el cruzamiento con ejemplares de otras poblaciones de galgos que se diferencien genéticamente para el carácter o caracteres de interés.
En estas notas nos vamos referir a la primera de las herramientas, la selección. Aparentemente estamos en una situación que se podría considerar muy favorable para aplicar selección genética utilizando métodos científicos, ya que existe una idea muy clara entre los criadores de que el componente genético que afecta a los principales caracteres en este tipo de galgos es muy relevante, como lo constata el hecho de que se paguen importantes cantidades de dinero por la utilización o adquisición como reproductores de ejemplares que han destacado en su comportamiento. Es decir, el criador de galgos considera que tanto un animal que ha tenido un buen comportamiento, como sus familiares estrechamente emparentados, son capaces de transmitir a su descendencia parte de esas buenas cualidades, y por ello están dispuestos a utilizar esos reproductores preferentemente, incluso a elevadosprecios. ¡¡Esto es la selección genética!!.
Si pudiéramos disponer de animales que corrían hace 100 años muchos quedarían sorprendidos de las enormes diferencias, no sólo morfológicas también de comportamiento, entre aquellos animales y los actuales (somos conscientes de que no se puede hablar de un único tipo de comportamiento, que éste ha podido variar mucho a lo largo del tiempo). Esos cambios entre aquellos animales y los actuales son básicamente consecuencia de la genética que subyace en la determinación, en mayor o menor medida, de todos esos caracteres.
¿De que manera podemos explotar la genética en esta población con el objetivo de mejorar aquellos caracteres que nos resultan de interés?
La selección genética
La seleccióngenética consiste en identificar a los animales portadores de los genes más beneficiosos para los caracteres de interés y utilizarlos como reproductores para que los transmitan a sus descendientes. El problema radica en que los animales no suelen llevar reflejado en ninguna parte de su exterior si es portador o no de genes favorables, y lo que tenemos que hacer es utilizar registros de algunas características (fenotipos) medidas (alzada, perímetro torácico, etc.) o valoradas en esos animales que puedan resultar un buen reflejo de la presencia o ausencia de esos genes que nos interesan. La forma de saber si la medida de un carácter (fenotipo) en un animal es o no un buen reflejo de los genes de que es portador ese animal (valor o mérito genético) es calculando la heredabilidad de ese carácter. Un valor elevado de la heredabilidad de un carácter nos indica que la medida de ese carácter en el propio animal, es decir, lo que se observa en un animal, nos proporciona una buena medida de la bondad de los genes de que es portador dicho animal, es decir, del mérito genético de ese animal. Así, un mal valor para un carácter de un animal nos está indicando que los genes que transmitirá a su descendencia no le permitirá a esta tener buenas medidas para ese carácter, y viceversa.
La heredabilidad
En primer lugar hay que entender que la heredabilidad de un carácter es un parámetro de la población en la que se registra dicho carácter, más adelante se pondrá algún ejemplo de lo que esto quiere decir. Entendido esto, la heredabilidad de un carácter es el cociente entre la variabilidad que observamos entre los méritos genéticos de los animales de una población para ese carácter (el numerador de la heredabilidad) y la variabilidad entre las medidas del carácter en los animales de la misma población (el denominador).
Si recordamos que el fenotipo, esto es, la medida que tenemos del carácter de interés, por ejemplo, cómo se ha comportado un animal, es el resultado de sumar al méritogenético el efecto de otro conjunto de factores que llamaremos nogenéticos, y que incluye cualquier factor que a uno se le ocurra no relacionado con la genética (año o época de nacimiento, tipo de nutrición, entrenamiento recibido, o la calidad de la propia medida del carácter, etc.), podemos darnos cuenta de que la heredabilidad de un carácter puede ser muy diferente de una población a otra, es decir, en una población de galgos un carácter puede manifestar un valor elevado de heredabilidad y ese mismo carácter un valor muy bajo en otra población. ¿Cuáles son las dos causas más importantes de que esto pueda ser así?
- Imaginemos dos poblaciones de galgos que tuvieran los mismo animales, es decir las dos fueran genéticamente idénticas, por lo tanto el numerador de la heredabilidad va a ser el mismo en ambas. Imaginemos también que en la primera población, para medir el peso al año, por ejemplo, (supongamos que es el carácter a mejorar) se utiliza una simple y vieja romana, mientras que en la segunda se utilizan básculas electrónicas con precisión de miligramo. En la primera población el peso de un animal al año va a ser medido con más error, posiblemente diferencias de hasta un kilogramo por medida (dependerá de lo bien calibrada que esté la romana), mientras que en la segunda el peso del animal va a ser registrado con absoluta precisión. En la primera población el denominador de la heredabilidad se va a ver incrementado respecto de la segunda precisamente por esta forma de medir con mucho error y, como el numerador de la heredabilidad en ambas poblaciones tiene el mismo valor ya que, como hemos indicado, tenían los mismos animales, el resultado es que la primera población, a pesar de disponer de la misma genética que la segunda, va a tener un valor de heredabilidad inferior al de la segunda.
- Supongamos que todos los animales de una población fueran genéticamente idénticos, por ejemplo ha incrementado tanto la consanguinidad que todos son genéticamente iguales. En esta situación no habría variabilidad entre los méritos genéticos de los animales de la población, todos los animales tendrían el mismo mérito genético, por lo tanto, el numerador de la heredabilidad sería nulo y nulo también el valor de la heredabilidad. En esta situación nos daría lo mismo seleccionar a los mejores o a los peores, genéticamente serían iguales e iguales serían en promedio sus descendencias.
¿Qué ocurre cuando el valor de la heredabilidad de un carácter es bajo (después comentaremos qué significa bajo o alto)?
En este caso la medida del propio animal (el fenotipo de ese animal) no resulta ser un buenreflejo de los genes de que es portador, es decir, puede ocurrir que animales con muy buen comportamiento tengan, con mucha frecuencia, descendientes con malos comportamientos, y viceversa. Con un valorbajo de heredabilidad, cometemos muchoserrores al elegir como bueno un animal del que sólo tenemos información de su comportamiento. En esta situación lo que tenemos que hacer es recabar más información sobre ese animal y ¿quién nos puede proporcionar más información sobre un animal?. En algunos casos el propio animal si el carácter que nos interesa puede medirse repetidas veces, pero sobre todo esa información adicional nos la puede proporcionar mediciones realizadas en parientes de ese animal, por ejemplo, las medidas que tengamos de sus padres, hermanos, medios hermanos o hijos. Por lo tanto, cuando un carácter manifiesta una heredabilidad suficientemente levada (por ejemplo, superior a 0,35 o 35%) las medidas de los caracteres en el propio individuo pueden proporcionarnos con suficiente precisión cual es el mérito genético de ese animal, si un carácter manifiesta una heredabilidad media (0,15-0,35) es conveniente utilizar información adicional de parientes. En el caso de valores de heredabilidad muy bajos (< 0,15) posiblemente la mejor estrategia de mejora no pase por la selección.
Métodos de selección
La información que nos proporciona sobre el valor genético de un animal una madre es equivalente a la que nos proporciona un hijo (en ambos casos está determinado por el parentesco entre madre e hijo), tengamos en cuenta, sin embargo, que un reproductor sólo tiene una madre, pero hijos puede tener muchos. Por esta razón se habla de selección por descendencia, cuando utilizamos la información de los numerosos hijos que puede tener un reproductor macho para conocer con precisión el mérito genético de ese macho, y selección por ascendencia, cuando utilizamos la información de los padres (o en general de ascendentes) para decidir sobre la bondad del mérito genético de un reproductor. En el caso de que sólo dispongamos de la información del propio animal, macho o hembra, para decidir si le seleccionamos o rechazamos como reproductor hablamos de selección individual.
La velocidad de selección
Es fácil entender que el progresogenético que se obtenga para un determinado carácter dependerá, por lo tanto, del valor de su heredabilidad, cuanto mayor sea esta mayor será el incremento que obtengamos en la media del carácter. El otro parámetro del que depende la ganancia genética que esperamos obtener es lo que podemos llamar la intensidad de selección. Esta intensidad será mayor cuanto mayor sea la diferencia entre los animales que hemos elegido como reproductores y la media de la población con la que se trabaja. Debemos darnos cuenta de que es este puede ser uno de los factores limitantes en el caso que nos ocupa. Esta diferencia entre lo que seleccionamos y la media que tenemos en nuestra población será mayor cuanto mayor sea el número de animales de los que tenemos registros de su comportamiento. Parece evidente que el mejor animal de 100 será, por término medio, peor que el mejor animal de 1.000 y mucho peor que el mejor animal de 1.000.000. Tengamos en cuenta que en la mejora genética en algunas especies de animales de renta, por ejemplo bovino lechero se utiliza información conjunta de millones de animales, se eligen las mejores vacas de entre cientos de miles de vacas y los mejores toros de entre miles de toros. Es muy probable que en el galgo de campo estas cifras resulten inalcanzables, lo cual repercute en la respuesta genética.
La ganancia genética que se obtiene depende, por lo tanto, del producto de la heredabilidad (h2) por la superioridad genética de los animales elegidos como reproductores (S):
R = h2 x S (1)
Y para expresar la ganancia genética en términos de velocidad de mejora, es decir, cual es la ganancia genética obtenida por año, tenemos que dividir R por el tiempo (t) que se tarda en obtener esa ganancia (igual que la velocidad es el espacio dividido por el tiempo):
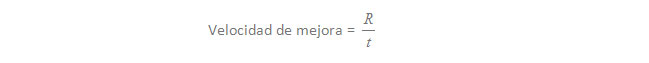
En nuestro contexto el tiempo se refiere al intervalo entre generaciones, es decir, la edad de un macho o de una hembra cuando nos deja un cachorro que elegimos para reproductor. La expresión (1) representa la ganancia genética esperada en una generación de selección, por lo tanto, para saber la velocidad de mejora debemos dividir esa ganancia por el tiempo que representa una generación de selección. Veamos un ejemplo práctico:
Supongamos que la media para un carácter de comportamiento en el galgo de campo es de 7 puntos en una escala de 1 a 10. Supongamos que los reproductores que elegimos tienen un valor medio de 9 puntos y que la heredabilidad de este carácter es de 0,35. Los hijos de esos reproductores, es decir, la generación siguiente estará por encima de la generación presente en aproximadamente:
0,35 * (9-7) = 0,7 puntos
es decir, el galgo de campo, fruto de la selección practicada tendrá ahora un valor medio de 7,7 puntos en ese carácter de comportamiento. Este avance se ha logrado en una generación de selección y cuanto más reducido sea éste mayor será la velocidad de mejora.
Un aspecto importante a destacar ahora es que este incremento puede parecer a primera vista poco importante, sin embargo hay que destacar que estos incrementos que se producen en cada generación son acumulativos y permanecen en la población aunque deje de practicarse selección. Por ello, los efectos de la selección deben observarse a medio o largo plazo y pueden resultar espectaculares si el plazo de tiempo que se contempla es elevado.
Al margen de algunos caracteres de tipo morfológico que pueden estar asociados a una buena presencia de los animales, los caracteres que parecen centrar la atención en la gran mayoría de los criadores de galgos son los relacionados con el comportamiento en los diferentes tipos de competiciones. La cuestión ahora sería, por lo tanto, responder a la pregunta sobre si practicar selección para estos caracteres nos permite abrigar esperanzas de que nuestra población de galgos se moverá en la dirección deseada por nosotros o no. ¿Qué podemos esperar?. Aunque completamente elucubrativo, si observamos lo que ocurre en otras especies, es muy probable que caracteres de comportamiento que, aunque puedan ser relativamente subjetivos, se midan concalidad en el galgo de campo es posible que el componentehereditario sea suficientemente elevado como para seleccionar con éxito sobre la base del mérito genético.
Pero para responder con precisión a la pregunta anterior es necesario calcular los valores de los principales parámetros genéticos de los caracteres de interés. Los dos parámetros genéticos relevantes serían las heredabilidades de los caracteres de interés, y las correlaciones genéticas entre dichos caracteres.
Del concepto y relevancia de la heredabilidad ya hemos hablado anteriormente, comentemos ahora brevemente el concepto y relevancia de las correlaciones genéticas entre caracteres.
La correlacióngenética puede ser consecuencia de varios fenómenos, pero el que nos interesa ahora es la correlación genética debida a un fenómeno que denominamos pleyotropía. La pleyotropía se refiere al hecho conocido de que un gen influye simultáneamente en la expresión de más de un carácter. Es fácil entender que existen muchos más fenotipos (caracteres) que genes por lo que es lógico esperar este fenómeno. Por ejemplo, es lógico pensar que algunos genes que influyen en el tamaño corporal también influyan en el peso del animal. Analizar las correlaciones genéticas es de gran interés ya que nos indica en que sentido se van a mover algunos caracteres por el hecho de modificar otros por selección. Es de gran interés saber si al seleccionar para mayor resistencia se pueden modificar en sentido no deseado otros caracteres, como velocidad inicial, etc. Por último, es importante no confundir la correlación fenotípica entre caracteres con la correlación genética. Nosotros podemos observar que existe correlación entre dos caracteres pero esto no significa en absoluto que esos cambios sean consecuencia genética. Es muy frecuente caer en ese error y argumentar que esas correlaciones fenotípicas que observamos entre dos caracteres algo tendrán de genético. La correlación entre dos variables no significa que una sea la causa de la otra, en muchas ocasiones existen otras variables no analizadas que son las que constituyen la causa de ambas y la causa de su correlación. Se suele citar un ejemplo muy ilustrativo de esto. Un estudio realizado en Inglaterra demostró la estrecha correlación entre el número de nidos de cigüeñas en un Condado y el número de nacimientos que se producían en ese Condado. Parecía que la conclusión era clara: los niños eran traídos por las cigüeñas. Evidentemente nuestro nivel de conocimientos nos dificultaba aceptar esta conclusión y la respuesta estaba precisamente en esa variable, antes comentada, que subyace a las otras dos (número de cigüeñas y número de niños nacidos) y que es la causa común que afecta a ambas variables, en este caso era el tamaño, la extensión, del Condado, cuanto mayor era el Condado mayor su número de habitantes, número de nacimientos y número de cigüeñas.
Por lo tanto, la única manera de saber si la modificación genética de un carácter llevará aparejada modificaciones genéticas en otros, el sentido en el que se modificarán, y de que magnitud en que lo harán es calculando las correlaciones genéticas entre esos caracteres. Dos caracteres con una correlación genética muy elevada (> 0,90), si en el que estamos interesados fuera difícil de medir o con baja heredabilidad frente al otro nos permitiría olvidarnos de él y seleccionar el otro.
Lo que nosotros observamos hoy en un carácter de un animal es el resultado tanto de la acción de los genes de que es portador dicho animal, como de un conjunto de factores no genéticos, como la forma de puntuar (es evidente que un mismo evaluador puede cambiar con el tiempo el criterio de puntuación de manera que lo que hace unos años lo veía como 7, hoy lo puntúa con un 5), el manejo, la alimentación, condiciones sanitarias, etc. Si nosotros pudiéramos separar los efectos genéticos de los no genéticos podríamos saber cual es el valor genético de cada uno de los animales de nuestra población pasada o presente. Si disponemos del mérito genético de cada uno de los animales a lo largo de los años podríamos calcular el valor medio de los méritos genéticos de todos los animales que han nacido durante un período de tiempo determinado, por ejemplo, durante un año. Si hacemos esto con todos los años de nuestra población tendremos una imagen de la tendencia genética a lo largo de toda la historia.
Los dosenemigos más generalizados para la práctica de la selección en este tipo de poblaciones son: a) La subjetividad de las puntuaciones que se asignan a los caracteres de comportamiento que puede llevar, si no se hacen con el acierto suficiente, a reducir drásticamente los valores de la heredabilidad, lo cual, a su vez, reduciría el progreso genético o lo haría prácticamente nulo; b) El reducidonúmero de reproductores por criador que puede dificultar la necesaria conexión genética entre las unidades de manejo.
Los métodos de valoracióngenética actuales permiten separar lo que es acción de genesde lo que son efectosnogenéticos. Así, una puntuación, por ejemplo, de 8 en un determinado carácter de comportamiento en un animal es explicada por un modelo que tiene en cuenta la acción del conjunto de genes de que es portador ese animal (lo que constituye el valor genético o mérito genético de ese animal), y un conjunto de factores no genéticos como, por ejemplo, el efecto del sexo del animal (si se trata de un macho o de una hembra), el efecto del criador, el efecto del año, en el que están incluidos no sólo efectos concretos de factores climáticos, también se incluyen aquí los avances en sanidad, alimentación, manejo y los cambios en la manera de puntuar para el carácter de comportamiento. Es fácil entender que dos animales con un mismo comportamiento de, por ejemplo, 8 pueden comportarse como reproductores de forma muy diferente, ya que en un caso el 8 de uno de los animales es fundamentalmente debido a los factores no genéticos, por lo que su descendencia no heredará los genes favorables, mientras que en el otro caso, el valor 8 de comportamiento es debido a los genes de que es portador y los factores no genéticos han aportado poco a ese comportamiento, en este caso la descendencia sí recibirá los genes favorables que le proporcionará un mejor comportamiento. Por lo tanto, dos animales con igual comportamiento no tienen la misma calidad como reproductores, esa calidad es lo que mide el mérito genético, que es el valor proporcionado por el modelo genético.
Actualmente la FEG tiene en proyecto informatizar toda la información disponible sobre el comportamiento de los galgos a lo largo de los últimos años. Con esta información no sólo se podrán conocer los valores de la heredabilidad y correlaciones entre caracteres, se podrá utilizar de forma simultánea para conocer la capacidad de transmisión a sus descendientes de un reproductor, es decir, se podrá disponer de un valor más preciso de un animal como reproductor, será posible disponer de una clasificación de los reproductores en función de su mérito genético para diferentes caracteres de comportamiento de tal forma que el criadorpodráseleccionar aquellos reproductores que mejor se acoplen a su situación y sus objetivos de mejora.